Superior colliculus
บทความนี้ต้องการตรวจสอบความถูกต้องจากผู้เชี่ยวชาญในเรื่องนั้น ๆ โปรดเพิ่มพารามิเตอร์ reason หรือ talk ลงในแม่แบบนี้เพื่ออธิบายปัญหาของบทความ |
บทความนี้มีชื่อเป็นภาษาอังกฤษ เนื่องจากชื่อเป็นศัพท์เฉพาะทาง ราชบัณฑิตยสถานยังไม่บัญญัติภาษาไทย หนังสือไทยใช้ศัพท์อังกฤษ |
optic tectum หรือเรียกสั้น ๆ ได้ว่า tectum เป็นโครงสร้างคู่ที่เป็นองค์ประกอบที่สำคัญในสมองส่วนกลางของสัตว์มีกระดูกสันหลัง ในสัตว์เลี้ยงลูกด้วยนม โครงสร้างนี้มักจะเรียกกันว่า superior colliculus[1] (ตัวย่อ SC) เป็นโครงสร้างที่มีลักษณะเป็นชั้น ๆ แม้ว่าจำนวนชั้นจะแตกต่างกันไปในสัตว์สปีชีส์ต่าง ๆ ชั้นนอก ๆ มีหน้าที่เกี่ยวกับประสาทสัมผัส และรับข้อมูลมาจากทั้งตาและระบบรับความรู้สึกอื่น ๆ [2] ส่วนชั้นที่ลึก ๆ ลงไปมีหน้าที่เกี่ยวกับการสั่งการ (motor) มีความสามารถในการเริ่มการเคลื่อนไหวของตาและเริ่มการตอบสนองในระบบอื่น ๆ ส่วนชั้นในระหว่างกลางมีนิวรอนที่ทำหน้าที่เกี่ยวกับประสาทสัมผัสหลายทาง และเกี่ยวกับการสั่งการด้วย
| Superior colliculus | |
|---|---|
 สมองตัดผ่าน superior colliculus (ไม่มีป้าย) แสดงเส้นประสาท oculomotor | |
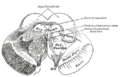
 แผนผังแสดงการเชื่อมต่อหลัก ๆ ของเส้นประสาทตา (optic nerves) และลำเส้นใยประสาทตา (optic tracts) ส่วน Superior colliculus อยู่ใกล้ศูนย์กลาง | |
| รายละเอียด | |
| ส่วนหนึ่งของ | เทคตัม |
| ตัวระบุ | |
| ภาษาละติน | Colliculus superior |
| MeSH | D013477 |
| นิวโรเนมส์ | 473 |
| นิวโรเล็กซ์ ID | birnlex_1040 |
| TA98 | A14.1.06.015 |
| TA2 | 5912 |
| TH | H3.11.03.3.01002 |
| TE | Terminologia Embryologica {{{2}}}.html EE5.14.3.3.1.4.4 .{{{2}}}{{{3}}} |
| FMA | 62403 |
| ศัพท์ทางกายวิภาคของประสาทกายวิภาคศาสตร์ | |
หน้าที่ทั่ว ๆ ไปของเทคตัมก็คือ ชี้ทางการตอบสนองทางพฤติกรรมไปยังตำแหน่งต่าง ๆ โดยมีกายเป็นศูนย์กลาง ชั้นแต่ละชั้นของเทตตัมมีแผนที่ภูมิลักษณ์ของโลกรอบตัวที่ใช้พิกัดแบบ retinotopy และการทำงานของนิวรอนจุดหนึ่งในแผนที่ทำให้เกิดการตอบสนองทางพฤติกรรมตรงตำแหน่งในปริภูมิที่สัมพันธ์กับจุดในแผนที่นั้น
ในไพรเมต งานศึกษาเรื่องของ SC โดยมากเป็นไปเกี่ยวกับการควบคุมการทอดสายตา. ข้อมูลทางตาที่มาจากจอตา หรือว่าสัญญาณสั่งการที่มาจากเปลือกสมอง สามารถเพิ่มระดับการทำงานของนิวรอนเป็นจุด ๆ ในแผนที่ ซึ่งถ้ามีกำลังพอ ก็จะก่อให้เกิดการเคลื่อนไหวตาแบบ saccade แต่ว่า แม้แต่ในไพรเมต SC ก็ยังมีหน้าที่เกี่ยวกับการหันศีรษะ การยื่นแขน[3] และการเปลี่ยนจุดที่ใส่ใจ โดยที่ไม่เกิดการเคลื่อนไหวที่เห็นได้[4] ในสปีชีส์อื่น ๆ เทคตัมมีหน้าที่มากมายรวมทั้ง การหันตัวในหนูที่กำลังเดิน ในปลาที่กำลังว่ายน้ำ ในนกที่กำลังบิน, การฉกเหยื่อด้วยลิ้นในกบ และการฉกเหยื่อในงูเป็นต้น
ในสปีชีส์อื่น ๆ นอกจากสัตว์เลี้ยงลูกด้วยนม รวมทั้งปลาและนก เทคตัมเป็นส่วนที่ใหญ่ที่สุดในสมอง ส่วนในสัตว์เลี้ยงลูกด้วยนมโดยเฉพาะในไพรเมต การขยายเปลือกสมองในลำดับวิวัฒนาการ ลดสัดส่วนของ SC เทียบกับสมองทั้งหมดลง แต่ถึงกระนั้น SC ก็ยังมีบทบาทที่สำคัญอย่างยิ่งโดยเป็นศูนย์ประสานงานหลักในการเคลื่อนไหวตา
เนื่องจากเอกสารหนังสือใช้ศัพท์ที่แตกต่างกันสำหรับสัตว์เลี้ยงลูกด้วยนมและสัตว์ประเภทอื่น ๆ สำหรับโครงสร้างที่จริง ๆ แล้วเป็นอันเดียวกัน นี่เป็นปัญหาสำหรับบทความที่จะอธิบายให้ครอบคลุมสปีชีส์ของสัตว์มีกระดูกสันหลังทั้งหมด ดูเหมือนว่า จะไม่มีวิธีอื่นที่จะอธิบายโดยไม่ก่อความรำคาญหรือความสับสนให้กับผู้อ่านบางท่าน บทความนี้จะใช้รูปแบบที่มีในเอกสารวิชาการต่าง ๆ คือใช้คำว่า superior colliculus เมื่อกล่าวถึงสัตว์เลี้ยงลูกด้วยนม และใช้คำว่า เทคตัม เมื่อกล่าวถึงสัตว์ประเภทอื่น
วิวัฒนาการและกายวิภาคเปรียบเทียบ
แก้
optic tectum เป็นส่วนที่สำคัญที่สุดส่วนหนึ่งในสมองของสัตว์มีกระดูกสันหลัง มีอยู่ในสปีชีส์ต่าง ๆ ตั้งแต่แฮคฟิชจนถึงมนุษย์[5] มีลักษณะบางอย่างที่เหมือนกันในสปีชีส์ต่าง ๆ รวมทั้งการแบ่งเป็นชั้น ๆ และมีแอกซอนส่งเข้ามาเป็นจำนวนมากจากลำเส้นใยประสาทตามายังชั้นนอก ๆ และจากระบบรับความรู้สึกทางกายมายังชั้นต่าง ๆ ด้านใน
ส่วนลักษณะอื่น ๆ มีความต่างกันไปในแต่ละสปีชีส์ เช่นจำนวนชั้นทั้งหมด (ตั้งแต่ 3 ในวงศ์ปลาปอดแอฟริกา ไปจนถึง 15 ในปลาทอง) [6] และประเภทต่าง ๆ ของเซลล์ (ตั้งแต่ 2 ประเภทในวงศ์ปลาปอดแอฟริกา จนถึง 27 ประเภทในนกกระจอกใหญ่) ในแฮคฟิช ปลาแลมป์เพรย์ทะเล และปลาฉลาม SC เป็นโครงสร้างที่เล็กเมื่อเทียบกับสมองส่วนอื่น ๆ แต่ในปลาที่มีก้านครีบ ที่มี infraclass เป็น "Teleostei" จะเป็นโครงสร้างที่ใหญ่ บางครั้งใหญ่ที่สุดในสมอง (ดูรูปสมองของปลาค็อดที่มีในบทความ) ในสัตว์สะเทินน้ำสะเทินบก สัตว์เลื้อยคลาน และโดยเฉพาะอย่างยิ่งสัตว์ปีก จะเป็นส่วนที่ค่อนข้างใหญ่ แต่ในสัตว์เลี้ยงลูกด้วยนม เป็นส่วนที่เล็กไปเมื่อเทียบกับเปลือกสมอง[6]
ปลาแลมป์เพรย์ทะเล
แก้มีการศึกษาปลาแลมป์เพรย์ทะเลกันอย่างกว้างขวางเพราะว่ามีสมองที่ค่อนข้างจะไม่ซับซ้อน ที่เชื่อกันว่า มีลักษณะหลายอย่างคล้ายกับโครงสร้างสมองของบรรพบุรุษสัตว์มีกระดูกสันหลัง ตั้งแต่คริสต์ทศวรรษ 1970 สเต็น กริวล์เนอร์และคณะที่สถาบัน Karolinska ในเมืองสต็อกโฮล์ม ได้ใช้ปลานี้เป็นตัวแบบเพื่อที่จะศึกษาหลักการควบคุมการเคลื่อนไหวของสัตว์มีกระดูกสันหลัง โดยเริ่มศึกษาตั้งแต่ไขสันหลังไปจนถึงสมอง[7]
ในผลงานวิจัยที่เรียงออกมาเป็นชุด พวกเขาได้พบว่า วงจรประสาทภายในไขสันหลัง มีความสามารถพอที่จะสร้างสัญญาณสั่งการเคลื่อนไหวที่มีรูปแบบเป็นจังหวะ (rhythmic) ซึ่งเป็นฐานของการว่ายน้ำ มีการควบคุมวงจรเหล่านี้โดยเขตประสาทที่ควบคุมการเคลื่อนไหวในก้านสมองและสมองส่วนกลาง ซึ่งก็มีการควบคุมอีกทีหนึ่งโดยเขตสมองต่าง ๆ ที่มีระดับสูงยิ่ง ๆ ขึ้นไปรวมทั้ง basal ganglia และเทคตัม
ในงานศึกษาเทคตัมโดยใช้ปลาแลมป์เพรย์ทะเลที่พิมพ์ในปี ค.ศ. 2007[8] พวกเขาพบว่า การกระตุ้นเทคตัมด้วยไฟฟ้าสามารถก่อให้เกิดการเคลื่อนไหวทางตา การงอตัวไปทางข้าง หรือการว่ายน้ำ และพบว่า ประเภท แอมพลิจูด และทิศทางของการเคลื่อนไหว มีความต่าง ๆ กันไป ขึ้นอยู่กับตำแหน่งของเทคตัมที่ได้รับการกระตุ้น หลักฐานนี้ ได้รับการพิจารณาว่า สอดคล้องกันความคิดว่า เทคตัมเป็นส่วนที่ก่อให้เกิดการเคลื่อนไหวที่มีเป้าหมายในทั้งปลาแลมป์เพรย์ทะเลและในสัตว์สปีชีส์อื่น ๆ
ค้างคาว
แก้ค้างคาวจริง ๆ แล้วไม่ได้เป็นสัตว์ตาบอด แต่ว่า เป็นสัตว์ที่อาศัยเสียงสะท้อนมากกว่าการมองเห็นในการบินไปในอากาศและในการจับเหยื่อ คือ ค้างคาวรับข้อมูลเกี่ยวกับโลกที่แวดล้อมโดยส่งเสียงร้องแหลมสั้น ๆ แล้วฟังเสียงสะท้อนกลับ สมองของค้างคาวมีการวิวัฒนาการในระดับสูงเพื่อทำกิจนี้โดยเฉพาะ และการปรับตัวเพื่อกิจนี้บางส่วนเกิดขึ้นที่ SC[9] ในค้างคาว เซลล์ที่รับข้อมูลจากจอตาเป็นชั้นบาง ๆ ใต้ผิวของ SC แต่มีส่วนที่ใหญ่กว้างขวางกว่าที่รับข้อมูลจากการได้ยิน และส่งแอกซอนไปยังเขตสั่งการ (motor area) ต่าง ๆ ที่สามารถให้เกิดการปรับทิศทางของหู หัว และร่างกาย เสียงสะท้อนที่มาจากทิศต่าง ๆ กันจะทำให้เกิดการตอบสนองในนิวรอนในตำแหน่งต่าง ๆ ในชั้นของ SC[10] และการทำงานของส่วนต่าง ๆ ของ SC ก็มีอิทธิพลต่อเสียงร้อง ดังนั้น นี้อาจจะเป็นหลักฐานที่ดีในทฤษฎีว่า SC มีส่วนในพฤติกรรมที่นำด้วยเสียงในค้างคาว เหมือนกับที่มีส่วนในพฤติกรรมที่นำด้วยการเห็นในสัตว์สปีชีส์อื่น ๆ
ค้างคาวมักจะจัดอยู่ในสองประเภท คือ อันดับย่อย Microchiroptera (ซึ่งมีมากที่สุด พบได้ทั่วโลก) และค้างคาวผลไม้ (พบได้ในเอเชีย แอฟริกา และออสเตรเลีย) แต่ค้างคาวผลไม้ไม่ได้กำหนดที่ตั้งวัตถุโดยเสียงสะท้อน แต่ใช้การเห็นเพื่อการนำทาง คือลานรับสัญญาณของนิวรอนใน SC เป็นแผนที่ภูมิลักษณ์ที่แม่นยำของเรตินา ซึ่งเหมือนกับที่พบในแมวและในไพรเมต
โครงสร้างและตำแหน่ง
แก้SC ที่มีเป็นคู่อยู่ใต้ทาลามัส รอบ ๆ ต่อมไพเนียลในสมองส่วนกลางของสัตว์มีกระดูกสันหลัง ประกอบด้วยส่วนด้านบน (dorsum) ของสมองส่วนกลาง หลัง periaqueductal gray และอยู่เหนือและติดกับ inferior colliculus โดยที่เมื่อรวม inferior colliculus กับ SC แล้ว จะเรียกโดยรวม ๆ กันว่า corpora quadrigemina (มาจากภาษาละติน แปลว่า ร่างมีสี่ส่วน)
วงจรประสาท
แก้

โครงสร้างละเอียดของ optic tectum หรือ SC มีความต่าง ๆ กันในแต่ละสปีชีส์ แต่โดยทั่ว ๆ ไปแล้ว ชั้นแรก ๆ ซึ่งรับข้อมูลจากระบบการมองเห็นและตอบสนองต่อข้อมูลทางตา จะมีความแตกต่างกันที่ชัดเจนจากชั้นต่าง ๆ ด้านใน ซึ่งรับข้อมูลมากมายหลายแบบและส่งข้อมูลไปยังเขตสั่งการ (motor) มากมายหลายเขตในสมอง ความแตกต่างระหว่างสองโซนนี้ชัดเจนและมีเหมือนกันในสัตว์สปีชีส์ต่าง ๆ จนกระทั่งว่า นักกายวิภาคบางพวกเสนอว่า ควรพิจารณาว่าเป็นโครงสร้างคนละส่วน
ในสัตว์เลี้ยงลูกด้วยนม นักกายวิภาคแบ่งโครงสร้างนี้ออกเป็น 7 ชั้น[12] 3 ชั้นแรกเรียกว่าชั้นใต้ผิว (superficial)
- ชั้นที่ 1 (stratum zonale ตัวย่อ SZ) เป็นชั้นบาง ๆ ประกอบด้วยแอกซอนมีปลอกไมอีลินเล็ก ๆ พร้อมกับเซลล์ marginal และ horizontal
- ชั้นที่ 2 (stratum griseum superficiale หรือ superficial gray ตัวย่อ SGS) มีนิวรอนที่มีรูปร่างและขนาดหลายอย่าง
- ชั้นที่ 3 (stratum opticum หรือ optic layer) โดยหลักประกอบด้วยแอกซอนจากลำเส้นใยประสาทตา
2 ชั้นต่อมาเรียกว่า ชั้นกลาง (intermediate)
- ชั้น 4 (stratum griseum intermediale หรือ intermediate gray) เป็นชั้นหนาที่สุด และมีนิวรอนมากมายหลายขนาด บ่อยครั้งหนาเท่าชั้นอื่น ๆ ทั้งหมดรวมกัน มักจะแบ่งย่อยเป็นส่วน "บน" และส่วน "ล่าง"
- ชั้น 5 (stratum album intermediale หรือ intermediate white) โดยมากเป็นใยประสาทมาจากที่ต่าง ๆ
2 ชั้นสุดท้ายเป็นชั้นที่อยู่ลึกที่สุด (deep)
- ชั้น 6 (stratum griseum profundum หรือ deep gray) เป็นชั้นที่อัดรวมกันอย่างหลวม ๆ ของนิวรอนและแอกซอนมีปลอกไมอีลิน
- ชั้น 7 (stratum album profundum หรือ deep white) อยู่ด้านบนและติดกับ periaqueductal gray โดยมากเป็นใยประสาท
ชั้นใต้ผิวรับข้อมูลโดยหลักจากจอตา จากเขตสายตาต่าง ๆ ในเปลือกสมอง และจากโครงสร้างที่เนื่องกับ SC ที่เรียกว่า pretectum และ parabigeminal nucleus ใยประสาทจากจอตาครอบคลุมทุกส่วนของชั้นใต้ผิว และเป็นใยประสาทที่มาจากตาทั้งสองข้าง แม้ว่าใยประสาทที่มาจากตาด้านตรงข้ามจะมีมากกว่า ส่วนใยประสาทจากคอร์เทกซ์มากที่สุดมาจากคอร์เทกซ์สายตาปฐมภูมิ (เขตบร็อดแมนน์ 17) คอร์เทกซ์สายตาทุติยภูมิ (เขตบร็อดแมนน์ 18 และเขตบร็อดแมนน์ 19) และจาก frontal eye fields ส่วน parabigeminal nucleus มีบทบาทที่สำคัญอย่างยิ่งในหน้าที่ของ SC ที่จะกล่าวต่อไป
เปรียบเทียบกับข้อมูลที่มาจากระบบสายตาของชั้นใต้ผิว ชั้นกลางและชั้นลึกรับข้อมูลโดยมากจากโครงสร้างของระบบรับความรู้สึกและระบบสั่งการ. เขตต่าง ๆ โดยมากจากเปลือกสมองจะส่งแอกซอนมายังชั้นต่าง ๆ เหล่านี้ แม้ว่า แอกซอนจากเขตสัมพันธ์ (association areas) มักจะหนาแน่นกว่าจากเขตรับความรู้สึกปฐมภูมิหรือจากเขตสั่งการ[ต้องการอ้างอิง] แต่จะมีความแตกต่างกันระหว่างสัตว์สปีชีส์ต่าง ๆ ทั้งในเขตเปลือกสมองที่ส่งข้อมูลมา ทั้งในความหนาแน่นของแอกซอนจากเขตนั้น ๆ [13]
แหล่งข้อมูลเข้าที่สำคัญอีกแหล่งหนึ่งมาจาก substantia nigra pars reticulata ซึ่งเป็นส่วนหนึ่งของ basal ganglia เป็นแหล่งข้อมูลแบบยับยั้ง (inhibitory) ที่ใช้สารสื่อประสาท GABA[14] ซึ่งเชื่อกันว่า มีส่วนในการช่วยควบคุม superior colliculus. ชั้นกลางและชั้นในรับข้อมูลจาก spinal trigeminal nucleus ซึ่งส่งข้อมูลความรู้สึกทางกาย (somatosensory) จากใบหน้า และจากไฮโปทาลามัส, zona incerta, ทาลามัส, และจาก inferior colliculus
นอกจากจะมีข้อมูลเข้าที่ต่าง ๆ กันแล้ว ชั้นใต้ผิวและชั้นลึกยังมีการส่งข้อมูลออกที่ต่าง ๆ กันอีกด้วย ข้อมูลที่สำคัญที่สุดส่วนหนึ่งจากชั้นใต้ผิวเดินทางไปยัง pulvinar และส่วนกลางด้านข้างของทาลามัส ซึ่งก็ส่งข้อมูลไปยังเปลือกสมองที่มีหน้าที่เกี่ยวข้องกับการควบคุมตาต่อไป และก็มีข้อมูลส่งจากชั้นใต้ผิวไปยัง pretectal nuclei, lateral geniculate nucleus ในทาลามัส, และ parabigeminal nucleus อีกด้วย แต่ข้อมูลที่ส่งไปจากชั้นลึกมีมากกว่า คือ มีวิถีประสาทขนาดใหญ่สองวิถีที่ส่งไปในระบบประสาทเบื้องต่ำ คือส่งไปยังก้านสมองและไขสันหลัง และยังมีวิถีประสาทมากมายส่งไปยังศูนย์รับความรู้สึก (sensory) และศูนย์สั่งการ (motor) รวมทั้งส่วนอื่น ๆ ที่มีส่วนร่วมในการสั่งการเคลื่อนไหวของตา
โครงสร้างที่แบ่งเป็นระบบย่อย ๆ
แก้ถ้าทำการสำรวจโดยละเอียดจะพบว่า ชั้นของ SC ไม่ได้เป็นแผ่นเรียบ ๆ แต่แบ่งออกเป็นคอลัมน์ต่าง ๆ จัดระเบียบคล้ายช่อง 6 เหลี่ยมในรวงผึ้ง[15] ตัวบ่งบอกถึงโครงสร้างเป็นคอลัมน์ที่ชัดเจนที่สุดมาจากแอกซอนที่ใช้สารสื่อประสาท acetylcholine จาก parabigeminal nucleus ที่มีส่วนสุด (terminal) เชื่อมกับ SC อย่างสม่ำเสมอตั้งแต่ด้านบนไปจนถึงด้านล่าง[16] สารเคมีประสาทที่เป็นตัวบ่งชี้อย่างอื่นรวมทั้ง calretinin, parvalbumin, GAP-43, และหน่วยรับความรู้สึกประเภท NMDA นอกจากนั้น การเชื่อมต่อกับโครงสร้างต่าง ๆ มากมายในก้านสมองและ diencephalon ก็ปรากฏลักษณะต่าง ๆ กันดังกล่าวนี้เช่นกัน[17] จำนวนคอลัมน์ทั้งหมดมีประมาณ 100[15] แม้ว่า ความสำคัญทางหน้าที่ของการจัดระเบียบเป็นคอลัมน์ยังไม่ชัดเจน แต่ก็น่าสนใจว่า หลักฐานที่ปรากฏเร็ว ๆ นี้แสดงว่า ใยประสาทที่ใช้สารสื่อประสาท acetylcholine เป็นส่วนของวงจรประสาทที่ทำให้เกิดความผันแปรแบบ winner-take-all ในเทคตัม ดังที่จะกล่าวต่อไป
ในสปีชีส์ที่ได้รับการสำรวจมาแล้วทั้งหมด รวมทั้งสัตว์เลี้ยงลูกด้วยนมและสัตว์อื่น ๆ มีการแบ่งออกเป็นส่วน ๆ เช่นนี้ แต่การแบ่งออกก็ยังมีความแตกต่างกันอย่างเป็นระบบในสปีชีส์ต่าง ๆ [16] คือ ในสปีชีส์ที่มีเรตินาแบบลาย (คือไม่มีรอยบุ๋มจอตาแต่มีแถบในจอตาที่เรียกว่า streak โดยหลัก ๆ แล้ว มีอยู่ในสปีชีส์ที่มีตาอยู่ทางด้านข้างเช่นกระต่ายและกวาง) การจัดระเบียบเช่นนี้ครอบคลุม SC ทั้งหมด แต่ในสปีชีส์ที่มีรอยบุ๋มจอตา (fovea) ที่อยู่ตรงกลาง การจัดระเบียบเช่นนี้ไปหยุดอยู่ที่ด้านหน้า (rostral) ของ SC ซึ่งเป็นส่วนที่มีนิวรอนมีหน้าที่เกี่ยวกับการตรึงตา (fixation) โดยยิงศักยะงานตลอดเวลาที่มีการตรึงตาในที่ใดที่หนึ่ง
Nucleus Isthmii/Parabigeminalis
แก้ส่วนนี้ไม่มีการอ้างอิงจากเอกสารอ้างอิงหรือแหล่งข้อมูล โปรดช่วยพัฒนาส่วนนี้โดยเพิ่มแหล่งข้อมูลน่าเชื่อถือ เนื้อหาที่ไม่มีการอ้างอิงอาจถูกคัดค้านหรือนำออก |

optic tectum มีการเชื่อมต่อกันอย่างสำคัญกับโครงสร้างที่ติดกันที่เรียกว่า nucleus isthmii ซึ่งดึงดูดความสนใจของนักวิจัยเร็ว ๆ นี้เพราะมีหลักฐานใหม่ ที่แสดงว่า nucleus isthmii มีความสำคัญในการทำหน้าที่ของเทคตัม แต่ในสัตว์เลี้ยงลูกด้วยนม ที่มีการใช้คำว่า superior colliculus มากกว่าที่จะใช้คำว่า optic tectum โครงสร้างนั้นกลับเรียกว่า parabigeminal nucleus นี่เป็นอีกส่วนหนึ่งในสมองที่มีการใช้ชื่อสองชื่อเรียกโครงสร้างอันเดียวกัน
nucleus isthmii นั้นแบ่งออกเป็นสองส่วน ส่วนแรกคือ pars magnocellularis (ตัวย่อ Imc แปลว่า ส่วนที่ประกอบด้วยเซลล์ขนาดใหญ่) และ pars parvocellularis (ตัวย่อ Ipc แปลว่า ส่วนที่ประกอบด้วยเซลล์ขนาดเล็ก) ส่วน Imc บางครั้งก็เรียกว่า pars semilunaris (ส่วนพระจันทร์ครึ่งดวง) เพราะมีรูปร่างคล้ายพระจันทร์ครึ่งดวง หรือเสี้ยวพระจันทร์ เมื่อผ่าออก
ดังที่แสดงในแผนผัง การเชื่อมต่อกันระหว่างเขต 3 เขตคือ เทคตัม, Ipc, และ Imc มีลักษณะเป็นแผนที่ภูมิลักษณ์ (topographic) คือ นิวรอนในชั้นนอก ๆ ของเทคตัมมีจุดที่สัมพันธ์กับนิวรอนของ Ipc และ Imc แอกซอนที่ส่งไปที่ Ipc มีการรวมศูนย์กันมากกว่าแอกซอนที่ส่งไปยัง Imc ซึ่งมีการกระจายออกในระดับที่สูงกว่าง ส่วน Ipc เองก็ส่งแอกซอนที่ใช้สารสื่อประสาท acetylcholine ไปยังทั้ง Imc และเทคตัม ในเทคตัม แอกซอนที่ใช้สารสื่อประสาท acetylcholine จาก Ipc แตกกิ่งก้านสาขาเดินทางไปไปยังจุดเป้าหมาย ที่ขยายไปทั่วทั้งคอลัมน์ ตั้งแต่บนจนถึงล่าง โดยเปรียบเทียบกัน Imc ส่งแอกซอนแบบ GABA[14] ไปยัง Ipc และเทคตัม ซึ่งกระจายไปในแนวด้านข้าง ครอบคลุมแผนที่ภูมิลักษณ์แบบ retinotopic ของส่วนทั้งสอง ดังนั้น จึงปรากฏว่า วงจรประสาท เทคตัม-Ipc-Imc ก่อให้เกิดการทำงานในเทคตัมเป็นการทำงานแบบมีสัญญาณป้อนกลับ ที่มีทั้งสัญญาณกระตุ้นที่รวมศูนย์จากคอลัมน์ของ Ipc และสัญญาณยับยั้งจากนิวรอนของ Imc ที่กระจายไปทั่ว
หน้าที่
แก้การศึกษาในเรื่อง optic tectum มีประวัติเกี่ยวกับการเปลี่ยนแนวความคิดหลายครั้งหลายคราว ก่อนปี ค.ศ. 1970 งานวิจัยโดยมากเป็นไปในสัตว์อื่นนอกจากสัตว์เลี้ยงลูกด้วยนม รวมทั้งปลา กบ และสัตว์ปีก คือเป็นการศึกษาในสปีชีส์ที่เทคตัมเป็นโครงสร้างหลักในการรับข้อมูลจากตา ความคิดพื้นฐานในเวลานั้นก็คือว่า ในสปีชีส์เหล่านี้ เทคตัมเป็นศูนย์การเห็นหลักในสมองของสัตว์อื่นที่ไม่ใช่สัตว์เลี้ยงลูกด้วยนม และดังนั้น จึงมีส่วนเกี่ยวข้องกับพฤติกรรมต่าง ๆ มากมาย
แต่ว่าในระหว่างคริสต์ทศวรรษ 1970 ถึง 1990 ก็มีการศึกษาด้วยการบันทึกสัญญาณประสาทในสัตว์เลี้ยงลูกด้วยนม เป็นไปโดยมากในลิง และโดยมากเพ่งจุดสนใจไปที่หน้าที่ของ superior colliculus ในการควบคุมการเคลื่อนไหวตา การศึกษาในแนวนี้รับการตีพิมพ์มากเสียจนกระทั่งว่า ความคิดของนักวิทยาศาสตร์โดยมากเห็นว่า การควบคุมตาเป็นหน้าที่สำคัญของ SC เพียงอย่างเดียวในสัตว์เลี้ยงลูกด้วยนม ซึ่งความเห็นเช่นนี้ก็ยังพบได้ในหนังสือวิชาการตราบเท่าทุกวันนี้
ในปลายคริสต์ทศวรรษ 1990 งานทดลองโดยใช้สัตว์ที่สามารถขยับหัวได้โดยอิสระ พบหลักฐานที่ชัดเจนว่า จริง ๆ แล้ว SC เป็นจุดกำเนิดการเปลี่ยนการทอดสายตา (gaze shifts) ซึ่งรวมทั้งการเคลื่อนศีรษะและการเคลื่อนตา คือไม่ใช่เป็นการเคลื่อนตาโดยอย่างเดียว การค้นพบนี้จุดประกายให้เกิดความสนใจในหน้าที่อย่างอื่น ๆ ของ SC อีก และนำไปสู่งานวิจัยที่พบการประมวลข้อมูลจากประสาทสัมผัสอื่น ๆ ในหลาย ๆ สปีชีส์ และในหลาย ๆ สถานการณ์ อย่างไรก็ดี บทบาทหน้าที่ของ SC ในการควบคุมตาเป็นเรื่องที่มีความเข้าใจดีกว่าหน้าที่อื่น ๆ ทั้งหมด
การศึกษาทางพฤติกรรมพบว่า SC ไม่จำเป็นในการรู้จำวัตถุ แต่มีบทบาทที่ขาดไม่ได้ในการชี้นำพฤติกรรมไปสู่วัตถุ ซึ่งสามารถเกิดขึ้นได้แม้จะขาดเปลือกสมอง[18] ดังนั้น แม้ว่า แมวที่มีความเสียหายออย่างสำคัญต่อคอร์เทกซ์สายตา (ในเปลือกสมอง) ไม่สามารถที่จะรู้จำวัตถุ แต่ก็อาจจะยังสามารถติดตามและขยับอวัยวะและตัวไปตามทิศทางของวัตถุที่เคลื่อนไหวอยู่ แม้ว่าอาจจะช้ากว่าปกติ แต่ถ้าว่า ครึ่งหนึ่งของ SC ถูกตัดออก แมวนั้นก็จะหมุนตัวไปรอบ ๆ ไปทางด้านของ SC ที่ถูกตัดออก และจะปรับอวัยวะและตัวเหมือนกับถูกบีบบังคับไปทางวัตถุที่อยู่ที่ด้านนั้น แต่จะไม่สามารถปรับอวัยวะและตัวไปสู่วัตถุที่อยู่ในด้านตรงกันข้าม ความบกพร่องอย่างนี้จะค่อย ๆ หายไปเมื่อเวลาผ่านไปแต่จะไม่หายไปโดยสิ้นเชิง
การเคลื่อนไหวตา
แก้ในไพรเมต การเคลื่อนไหวตานั้นสามารถแบ่งได้เป็น 4 อย่าง คือ
- การตรึงตา (fixation) เป็นการตรึงตาทั้งสองที่วัตถุที่อยู่นิ่ง ๆ โดยจะมีการเคลื่อนไหวตาเพื่อชดเชยการเคลื่อนไหวของศีรษะเท่านั้น
- smooth pursuit เป็นการเคลื่อนไหวตาอย่างสม่ำเสมอตามวัตถุที่กำลังเคลื่อนที่
- saccades เป็นการเคลื่อนไหวตาอย่างรวดเร็วจากตำแหน่งหนึ่งไปยังอีกตำแหน่งหนึ่ง
- vergence เป็นการเคลื่อนไหวตาทั้งสองข้างไปยังทิศทางตรงกันข้ามพร้อม ๆ กันเพื่อที่จะได้มาหรือดำรงไว้ซึ่งการเห็นแบบ binocular
แม้ว่า SC จะมีบทบาทในการเคลื่อนไหวตาเหล่านี้ทั้งหมด แต่หน้าที่เกี่ยวกับ saccades เป็นส่วนที่ได้รับการศึกษาแล้วมากที่สุด
SC แต่ละข้างของซีกสมอง มีแผนที่ 2 มิติเป็นตัวแทนของลานสายตากึ่งด้าน ส่วนรอยบุ๋มจอตา (fovea) ซึ่งเป็นส่วนที่สามารถรับข้อมูลได้อย่างละเอียดที่สุด ปรากฏที่สุดด้านหน้าของแผนที่ และส่วนรอบ ๆ สายตา (peripheral) ปรากฏที่สุดด้านหลังของแผนที่ การเคลื่อนไหวตาเกิดขึ้นเพราะการทำงานในชั้นลึก ๆ ของ SC
ในระหว่างการตรึงตา นิวรอนใกล้ด้านหน้า ซึ่งรับข้อมูลมาจากรอยบุ๋มจอตา จะมีการส่งสัญญาณแบบ tonic (ไปเรื่อย ๆ แบบไม่ถี่) ส่วนในระหว่าง smooth pursuit นิวรอนส่วนกระเถิบไปอีกหน่อยหนึ่งจากด้านหน้าจะส่งสัญญาณ ซึ่งนำไปสู่การเคลื่อนไหวตาแบบเป็นไปทีละน้อย สำหรับ saccades นิวรอนซึ่งเป็นตัวแทนจุดที่ตาจะไปทอดลง จะเกิดการทำงาน คือ ก่อนที่ saccade จะเกิดขึ้น จะมีการเพิ่มการทำงานของนิวรอนในจุดที่ตาจะไปทอดลงอย่างรวดเร็ว และการทำงานในส่วนอื่น ๆ ของ SC ก็จะลดลง แต่ว่า การเข้ารหัสตำแหน่งค่อนข้างที่จะกว้าง คือ สำหรับการเคลื่อนไหวตาแบบ saccade ครั้งหนึ่ง ๆ ระดับการส่งสัญญาณของนิวรอนรอบ ๆ จะมีลักษณะเป็น "เนินเขา" ซึ่งครอบคลุมส่วนหนึ่ง ๆ ของแผนที่ใน SC โดยที่ตำแหน่งของ "ยอดเขา" เป็นตำแหน่งเป้าหมายของ saccade
แม้ว่า SC จะเข้ารหัสจุดเป้าหมายของการเปลี่ยนการทอดสายตา แต่ไม่ปรากฏว่าเป็นส่วนที่กำหนดการเคลื่อนไหวตาและศีรษะเป็นลำดับเพื่อจะให้สำเร็จเป้าหมายนั้นได้[19] การแปลงสัญญาณเป้าหมายการทอดสายตาออกให้เป็นการเคลื่อนไหวศีรษะและตา ให้เป็นวิถีการเคลื่อนไหวของตาในระหว่าง saccade อาศัยการประมวลผลข้อมูลจาก SC และจากส่วนอื่น ๆ ในสมอง โดยเขตสั่งการต่าง ๆ ที่รับสัญญาณจาก SC ซึ่งเป็นกระบวนการที่ยังไม่มีความเข้าใจกันดี แต่ไม่ว่ากระบวนการนั้นจะเป็นไปได้อย่างไร SC ก็ยังเป็นส่วนที่เข้ารหัสตำแหน่งเป้าหมายโดยแผนที่ภูมิลักษณ์แบบ retinotopic นั่นก็คือ รูปแบบการทำงานของ SC บอกถึงออฟเซต (ระยะและทิศทางที่แตกต่าง) จากตำแหน่งที่ตากำลังจ้องมอง โดยไม่เกี่ยวข้องว่าตำแหน่งเริ่มต้นของตาจะอยู่ที่ไหน[20]
ยังมีข้อขัดแย้งว่า SC เพียงแต่เริ่มสั่งการเคลื่อนไหวของตา แล้วโครงสร้างอื่น ๆ จึงทำการปฏิบัติการ หรือว่า แม้ SC ก็ทำการปฏิบัติการด้วยเกี่ยวกับการเคลื่อนไหวตาแบบ saccade ในปี ค.ศ. 1991 มูโนซ์และคณะ เสนอโดยใช้ข้อมูลจากงานวิจัยของตนว่า ในช่วงที่เกิด saccade "เนินเขา" ที่แสดงการทำงานของ SC มีการเคลื่อนไปอย่างช้า ๆ ซึ่งแสดงออฟเซตที่กำลังแปรเปลี่ยนไปของตาจากตำแหน่งเป้าหมาย[21] ถึงอย่างนั้น ในปัจจุบัน ความเห็นหลักของเหล่านักวิทยาศาตร์ก็คือว่า แม้ว่า เนินเขานั้นจะย้ายตำแหน่งไปเล็ก ๆ น้อย ๆ ในระหว่าง saccade แต่ว่า การย้ายตำแหน่งนั้นไม่เป็นไปอย่างคงที่คงวาและไม่เป็นสัดส่วนดังที่สมมุติฐาน "เนินเขาเคลื่อนที่" นั้นพยากรณ์[22]
ส่วนสั่งการของ SC ส่งสัญญาณขาออกไปยังนิวเคลียสในสมองส่วนกลางและก้านสมอง ซึ่งเปลี่ยนการเข้ารหัสโดยตำแหน่งใน SC ไปเป็นการเข้ารหัสโดยความถี่ศักยะงานที่ใช้กันในกลุ่มนิวรอน oculomotor การเคลื่อนไหวตาเกิดขึ้นได้อาศัยกล้ามเนื้อหกมัด ซึ่งแบ่งเป็น 3 คู่ คู่หนึ่ง ๆ ตั้งฉากกับคู่อื่น ๆ ดังนั้น ในระดับสุดท้ายของการเคลื่อนไหวตา การเข้ารหัสเป็นไปตามระบบพิกัดคาร์ทีเซียน
แม้ว่า SC จะรับสัญญาณโดยตรงที่มีกำลังจากจอตา แต่ในไพรเมต SC จะได้รับการควบคุมจากเปลือกสมองโดยหลัก ซึ่งประกอบด้วยเขตต่าง ๆ ที่มีหน้าที่กำหนดการเคลื่อนไหวตา[23] คือ
- frontal eye fields (FEF) ซึ่งเป็นส่วนของคอร์เทกซ์สั่งการ มีหน้าที่เริ่มต้น saccades ที่อยู่ใต้อำนาจจิตใจ
- supplementary eye fields ซึ่งอยู่ติดกับ FEF มีหน้าที่แบ่งการเคลื่อนไหวแบบ saccades ออกเป็นลำดับ
- parietal eye fields ซึ่งอยู่เลยไปทางด้านหลังของสมอง มีหน้าที่โดยหลักเกี่ยวกับการเคลื่อนไหวแบบ saccades ที่เป็นรีเฟล็กซ์ ซึ่งเกิดขึ้นตอบสนองการเปลี่ยนแปลงบางอย่างของวิวที่เห็น
SC รับข้อมูลจากตาที่ชั้นนอก ๆ เท่านั้น คือชั้นที่ลึกลงไปรับข้อมูลจากหูและจากกาย และมีการเชื่อมต่อกับเขตสั่งการที่ตอบสนองต่อข้อมูลประสาทสัมผัสต่าง ๆ โดยรวม ๆ แล้ว เชื่อกันว่า SC มีหน้าที่ช่วยปรับศีรษะและตาไปทางสิ่งที่เห็นและได้ยิน[4][24][25][26]
SC รับข้อมูลการได้ยินมาจาก inferior colliculus มีการประสานข้อมูลทางหูกับข้อมูลทางตามีผลเป็นการรับรู้ว่า ใครเป็นคนพูด (และทำให้เกิดการแปลสิ่งเร้าผิดว่า หุ่นที่อยู่ในมือเป็นผู้พูดแทนผู้พากย์เสียง)

ความหลากหลาย
แก้ไพรเมต
แก้เป็นที่ยอมรับกันว่า SC ในไพรเมตไม่เหมือนกับในสัตว์เลี้ยงลูกด้วยนมประเภทอื่น ๆ เพราะว่าไม่มีแผนที่ของลานสายตาทั้งหมดที่มาจากตาด้านตรงกันข้าม แต่ว่า โดยเหมือนกับคอร์เทกซ์สายตาและ lateral geniculate nucleus SC มีแผนที่ครึ่งหนึ่งของลานสายตาจากตาทั้งสอง (คือ SC ในสมองซีกซ้ายมีแผนที่ลานสายตาด้านขวาทั้งจากตาซ้ายและตาขวา) แต่ไม่มีแผนที่อีกครึ่งหนึ่งของลานสายตาด้านเดียวกัน (จากตาทั้งสอง) [27]
ที่เป็นเช่นนี้ก็เพราะว่า ไพรเมตไม่มีการเชื่อมต่อกันระหว่าง retinal ganglion cell ในกึ่งข้างใน (ข้างขมับ) ของจอตา กับ SC ซีกตรงกันข้าม ในสัตว์เลี้ยงลูกด้วยนมประเภทอื่น retinal ganglion cell ในจอตา ส่งแอกซอนทั้งหมดไปยัง SC ซีกตรงกันข้าม
ความแตกต่างเช่นนี้ระหว่างไพรเมตกับสัตว์เลี้ยงลูกด้วยนมประเภทอื่น เป็นหลักฐานชิ้นสำคัญของทฤษฎีว่า ค้างคาวผลไม้มีสัตว์บรรพบุรุษเดียวกัน (sister taxon) กับไพรเมต ซึ่งเป็นทฤษฏีเสนอโดยนักประสาทวิทยาศาสตร์ชาวออสเตรเลีย ดร. แจ็ค เพ็ตติกริว ในปี ค.ศ. 1986 หลังจากที่ได้พบว่า ค้างคาวผลไม้มีความคล้ายคลึงกับไพรเมต โดยมีการเชื่อมต่อกันระหว่างจอตาและ SC ที่เหมือนกัน ซึ่งต่างจากสัตว์เลี้ยงลูกด้วยนมประเภทอื่น ๆ [28]
สัตว์มีกระดูกสันเหลังประเภทอื่น ๆ
แก้ในตระกูลงูที่สามารถตรวจจับรังสีอินฟราเรดได้ เช่นวงศ์งูเหลือมและวงศ์ย่อยงูหางกระดิ่ง เส้นประสาทจากตาส่งผ่าน trigeminal nerve แทนที่จะผ่านลำเส้นใยประสาทตา แต่ว่า การแปลผลข้อมูลอย่างอื่นเหมือนกัน ดังนั้น งูเหล่านั้นจึงมี optic tectum ด้วยเช่นกัน[29]
ดูเพิ่ม
แก้รูปภาพอื่น ๆ
แก้-
 ก้านสมอง ชำแหละโดยตื้น มองจากด้านข้าง (lateral)
ก้านสมอง ชำแหละโดยตื้น มองจากด้านข้าง (lateral) -
 ก้านสมอง มองจากด้านข้าง (lateral)
ก้านสมอง มองจากด้านข้าง (lateral) -
 ก้านสมอง ชำแหละโดยลึก มองจากด้านข้าง (lateral)
ก้านสมอง ชำแหละโดยลึก มองจากด้านข้าง (lateral) -
 ก้านสมอง ชำแหละโดยลึก มองจากด้านข้าง (lateral)
ก้านสมอง ชำแหละโดยลึก มองจากด้านข้าง (lateral) -
 สมองส่วนกลาง ผ่าตามขวาง (Transverse) ที่ระดับของ SC
สมองส่วนกลาง ผ่าตามขวาง (Transverse) ที่ระดับของ SC -

-
 Superior colliculus
Superior colliculus







แหล่งข้อมูลอื่น
แก้- ก้านสมองที่มหาวิทยาลัยวิสคอนซิน 23Colliculus
- ภาพสมองตัดแต่งสีซึ่งรวมส่วน "superior colliculus" at the BrainMaps project
เชิงอรรถและอ้างอิง
แก้- ↑ ศ.พญ. ผาสุก มหรรฆานุเคราะห์ (พ.ศ. 2556). ประสาทกายวิภาคศาสตร์พื้นฐาน (ฺBasic neuroanatomy). กรุงเทพมหานคร: ศ.พญ. ผาสุก มหรรฆานุเคราะห์. p. 258. ISBN 978-616-335-105-0.
{{cite book}}: ตรวจสอบค่าวันที่ใน:|year=(help) - ↑ Wallace et al., 2005
- ↑ Lunenburger et al., 2001
- ↑ 4.0 4.1 Kustov & Robinson, 1996
- ↑ Maximino, 2008
- ↑ 6.0 6.1 Northcutt, 2002
- ↑ Grillner, 2003
- ↑ Saitoh et al., 2007
- ↑ Ulanovsky & Moss, 2008
- ↑ Valentine & Moss, 1997
- ↑ Caltharp SA, Pira CU, Mishima N, Youngdale EN, McNeill DS, Liwnicz BH, Oberg KC (2007). "NOGO-A induction and localization during chick brain development indicate a role disparate from neurite outgrowth inhibition". BMC Dev. Biol. 7 (1): 32. doi:10.1186/1471-213X-7-32. PMC 1865376. PMID 17433109.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ Huerta & Harting, 1984
- ↑ Clemo HR, Stein BE (1984). "Topographic organization of somatosensory corticotectal influences in cat". Journal of Neurophysiology. 51 (5): 843–858. PMID 6726314. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2008-07-03. สืบค้นเมื่อ 2014-06-07.
- ↑ 14.0 14.1 กาบา (Gamma-Aminobutyric acid ตัวย่อ GABA) เป็นสารสื่อประสาทแบบห้ามตัวหลัก ในระบบประสาทกลางของสัตว์เลี้ยงลูกด้วยนม มีบทบาทสำคัญในการควบคุมความกระตุ้นได้โดยทั่วไปในระบบประสาท นอกจากนั้นแล้ว ในมนุษย์ กาบายังมีบทบาทโดยตรงในการควบคุมความตึงตัวของกล้ามเนื้อ
- ↑ 15.0 15.1 Chavalier & Mana, 2000
- ↑ 16.0 16.1 Illing, 1996
- ↑ Mana & Chevalier, 2001
- ↑ Sprague, 1996
- ↑ Sparks & Gandhi, 2003
- ↑ Klier et al., 2001
- ↑ Munoz et al., 1991
- ↑ Soetedjo et al., 2002
- ↑ Pierrot-Deseilligny et al., 2003
- ↑ Klier et al., 2003
- ↑ Krauzlis et al., 2004
- ↑ Sparks, 1999
- ↑ Lane et al., 1973
- ↑ Pettigrew, 1986
- ↑ Hartline et al., 1978
อ้างอิง
แก้- Chevalier, G; Mana S (2000). "Honeycomb-like structure of the intermediate layers of the rat superior colliculus, with additional observations in several other mammals: AChE patterning". J Comp Neurol. 419 (2): 137–53. doi:10.1002/(SICI)1096-9861(20000403)419:2<137::AID-CNE1>3.0.CO;2-6. PMID 10722995. S2CID 26050145.
- Dash, S; Yang X; Wang H; Crawford JD (2015). "Continuous updating of visuospatial memory in superior colliculus during slow eye movements". Curr Biol. 25 (3): 267–74. doi:10.1016/j.cub.2014.11.064. PMID 25601549.
- Dean, P; Redgrave P; Westby GW (1989). "Event or emergency? Two response systems in the mammalian superior colliculus". Trends Neurosci. 12 (4): 137–47. doi:10.1016/0166-2236(89)90052-0. PMID 2470171. S2CID 25268593.
- Gandhi, NJ; Katani HA (2011). "Motor Functions of the Superior Colliculus". Annu Rev Neurosci. 34: 205–231. doi:10.1146/annurev-neuro-061010-113728. PMC 3641825. PMID 21456962.
- Grillner, S (2003). "The motor infrastructure: from ion channels to neuronal networks". Nature Reviews Neuroscience. 4 (7): 573–86. doi:10.1038/nrn1137. PMID 12838332. S2CID 4303607.
- Hartline, PH; Kass L; Loop MS (1978). "Merging of modalities in the optic tectum: infrared and visual integration in rattlesnakes". Science. 199 (4334): 1225–9. Bibcode:1978Sci...199.1225H. doi:10.1126/science.628839. PMID 628839.
- Huerta, MF; Harting JK (1984). Vanegas H (บ.ก.). Comparative Neurology of the Optic Tectum. New York: Plenum Press. pp. 687–773. ISBN 978-0-306-41236-3.
- Illing, R-B (1996). "Chapter 2 the mosaic architecture of the superior colliculus". Extrageniculostriate Mechanisms Underlying Visually-Guided Orientation Behavior. Prog Brain Res. Progress in Brain Research. Vol. 112. pp. 17–34. doi:10.1016/S0079-6123(08)63318-X. ISBN 9780444823472. PMID 8979818.
- King, AJ; Schnupp JWH; Carlile S; Smith AL; Thompson ID (1996). "Chapter 24 the development of topographically-aligned maps of visual and auditory space in the superior colliculus". Extrageniculostriate Mechanisms Underlying Visually-Guided Orientation Behavior. Prog Brain Res. Progress in Brain Research. Vol. 112. pp. 335–350. doi:10.1016/S0079-6123(08)63340-3. ISBN 9780444823472. PMID 8979840.
- Klier, EM; Wang H; Crawford JD (2001). "The superior colliculus encodes gaze commands in retinal coordinates" (PDF). Nat Neurosci. 4 (6): 627–32. doi:10.1038/88450. PMID 11369944. S2CID 4930662.
- Klier, E; Wang H; Crawford D (2003). "Three-dimensional eye-head coordination is implemented downstream from the superior colliculus". J Neurophysiol. 89 (5): 2839–53. CiteSeerX 10.1.1.548.1312. doi:10.1152/jn.00763.2002. PMID 12740415.
- Krauzlis, R; Liston D; Carello C (2004). "Target selection and the superior colliculus: goals, choices and hypotheses". Vision Res. 44 (12): 1445–51. doi:10.1016/j.visres.2004.01.005. PMID 15066403. S2CID 5705421.
- Kustov, A; Robinson D (1996). "Shared neural control of attentional shifts and eye movements". Nature. 384 (6604): 74–77. Bibcode:1996Natur.384...74K. doi:10.1038/384074a0. PMID 8900281. S2CID 68917.
- Lane, RH; Allman JM; Kaas JH; Miezin FM (1973). "The visuotopic organization of the superior colliculus of the owl monkey (Aotus trivirgatus) and the bush baby (Galago senegalensis)". Brain Res. 60 (2): 335–49. doi:10.1016/0006-8993(73)90794-4. PMID 4202853.
- Lunenburger, L; Kleiser R; Stuphorn V; Miller LE; Hoffmann KP (2001). "Chapter 8 a possible role of the superior colliculus in eye-hand coordination". Vision: From Neurons to Cognition. Prog Brain Res. Progress in Brain Research. Vol. 134. pp. 109–25. doi:10.1016/S0079-6123(01)34009-8. ISBN 9780444505866. PMID 11702538.
- Mana, S; Chevalier G (2001). "Honeycomb-like structure of the intermediate layers of the rat superior colliculus: afferent and efferent connections". Neuroscience. 103 (3): 673–93. doi:10.1016/S0306-4522(01)00026-4. PMID 11274787. S2CID 45660874.
- Maximino, C; Soares, Daphne (2008). Soares, Daphne (บ.ก.). "Evolutionary changes in the complexity of the tectum of nontetrapods: a cladistic approach". PLOS ONE. 3 (10): e385. Bibcode:2008PLoSO...3.3582M. doi:10.1371/journal.pone.0003582. PMC 2571994. PMID 18974789.
- Munoz, DP; Pélisson D; Guitton D (1991). "Movement of activity on the superior colliculus motor map during gaze shifts" (PDF). Science. 251 (4999): 1358–60. doi:10.1126/science.2003221. PMID 2003221.
- Northcutt, RG (2002). "Understanding vertebrate brain evolution". Integr Comp Biol. 42 (4): 743–6. doi:10.1093/icb/42.4.743. PMID 21708771.
- Pettigrew, JD (1986). "Flying primates? Megabats have the advanced pathway from eye to midbrain". Science. 231 (4743): 1304–6. Bibcode:1986Sci...231.1304P. doi:10.1126/science.3945827. PMID 3945827. S2CID 16582493.
- Pierrot-Deseilligny, C; Müri RM; Ploner CJ; Gaymard B; Rivaud-Péchoux S (2003). "Cortical control of ocular saccades in humans: A model for motricity". Neural Control of Space Coding and Action Production. Prog Brain Res. Progress in Brain Research. Vol. 142. pp. 3–17. doi:10.1016/S0079-6123(03)42003-7. ISBN 9780444509772. PMID 12693251.
- Saitoh, K; Ménard A; Grillner S (2007). "Tectal control of locomotion, steering, and eye movements in lamprey". J Neurophysiol. 97 (4): 3093–108. doi:10.1152/jn.00639.2006. PMID 17303814. S2CID 5711513.
- Soetedjo, R; Kaneko CR; Fuchs AF (2002). "Evidence against a moving hill in the superior colliculus during saccadic eye movements in the monkey". J Neurophysiol. 87 (6): 2778–89. doi:10.1152/jn.2002.87.6.2778. PMID 12037180. S2CID 18294502.
- Sparks, DL (1999). "Conceptual issues related to the role of the superior colliculus in the control of gaze". Current Opinion in Neurobiology. 9 (6): 698–707. doi:10.1016/S0959-4388(99)00039-2. PMID 10607648. S2CID 14389002.
- Sparks, DL; Gandhi NJ (2003). "Single cell signals: An oculomotor perspective". Neural Control of Space Coding and Action Production. Prog Brain Res. Progress in Brain Research. Vol. 142. pp. 35–53. doi:10.1016/S0079-6123(03)42005-0. ISBN 9780444509772. PMID 12693253.
- Sprague, JM (1996). "Chapter 1 Neural mechanisms of visual orienting responses". Extrageniculostriate Mechanisms Underlying Visually-Guided Orientation Behavior. Prog Brain Res. Progress in Brain Research. Vol. 112. pp. 1–15. doi:10.1016/S0079-6123(08)63317-8. ISBN 9780444823472. PMID 8979817.
- Stein, BE; Clamman HP (1981). "Control of pinna movements and sensorimotor register in cat superior colliculus". Brain Behav Evol. 19 (3–4): 180–192. doi:10.1159/000121641. PMID 7326575.
- Ulanovsky, N; Moss CF (2008). "What the bat's voice tells the bat's brain". PNAS. 105 (25): 8491–98. Bibcode:2008PNAS..105.8491U. doi:10.1073/pnas.0703550105. PMC 2438418. PMID 18562301.
- Valentine, D; Moss CF (1997). "Spatially selective auditory responses in the superior colliculus of the echolocating bat". J Neurosci. 17 (5): 1720–33. doi:10.1523/JNEUROSCI.17-05-01720.1997. PMC 6573370. PMID 9030631.
- Wallace, MT; Meredith MA; Stein BE (1998). "Multisensory integration in the superior colliculus of the alert cat". J Neurophysiol. 80 (2): 1006–10. doi:10.1152/jn.1998.80.2.1006. PMID 9705489.